Biokémia
← ElőzőOldal 4 / 4
Jelentkezz be a hozzászóláshoz.

#10
köszönöm szépen!😊 kurva érdekes... azthiszem most egy darabig maradok is read-only módban, csak hajrá! érdekes téma, jó olvasgatni ilyeneket is

#9
ugyanez van az aminosavakkal is, az α-C-atom királis, létezik tükörképi párja, éppen ezt raktam be az elsõ képre.
Amúgy ezt sztereokémiának hívják.
Az enzimek általában szterospecifikusak, tehát pl csak a D-glükózt tudják lebontani, ezért a természetben általában csak D cukrok és L aminosavak vannak.
Amúgy ezt sztereokémiának hívják.
Az enzimek általában szterospecifikusak, tehát pl csak a D-glükózt tudják lebontani, ezért a természetben általában csak D cukrok és L aminosavak vannak.
dixitque deus fiat lux
#8
másik kép...
dixitque deus fiat lux
#7
igen, ez így van.
Onnan érdemes megközelíteni a dolgot, hogy pl. a glükóz egy monoszacharid, ami azt jelenti, hogy 1 cukormolekula, van diszacharid(2 molek),oligoszacharid(10-es nagyságrend),poliszacharid(100-as és azon felüli nagyságrend) is.
Még az is köztudott, hogy szénhidrátnak nevezzük. Ez az elnevezés onnan jön, hogy régen a molekula szerkezetét teljesen nem ismerve, azt hitték, hogy C
Tehát ha megnézzük akkor C,O,H-bõl áll, és amit írtál, hogy a cukrokból többféle létezik a C miatt van.
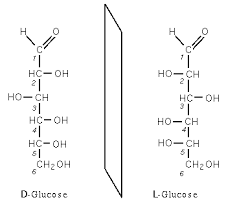
Ha van egy olyan C atom, amihez kapcsolódik 4 különbözõ másik ligandum (atom v molekula), akkor ebbõl a C atomból 2 féle létezik, melyek egymásnak tükörképei (királis), mint a két tenyér:

Ez csak akkor történik meg, ha a C-hez 4 különbözõ szubsztituens kapcsolódik, ha ezek közül 2 egyforma, akkor úgy összeforgatható a két C, hogy azok egymást fedik, tehát nem tükörképek.
A cukrokban is van ilyen C-atom amelyik királis:
http://t2.gstatic.com/images?q=tbn:7MKGzF1J4GvWLM:http://fr.wikivisual.com/images/5/53/D_et_l_glucose.png&t=1
Amint a képbõl is látszik, egymás tükörképe a két molekula, ugyanolyan atomok alkotják, ugyanolyan kapcsolódási sorrendben, de más térszerkezetben.Az egyiket D (dexter-jobb) a másikat L (leveus-bal) glükóznak nevezik, az alapján, hogy a királis C-atom az ún. Fischer-projekciós ábrán merre áll.
Onnan érdemes megközelíteni a dolgot, hogy pl. a glükóz egy monoszacharid, ami azt jelenti, hogy 1 cukormolekula, van diszacharid(2 molek),oligoszacharid(10-es nagyságrend),poliszacharid(100-as és azon felüli nagyságrend) is.
Még az is köztudott, hogy szénhidrátnak nevezzük. Ez az elnevezés onnan jön, hogy régen a molekula szerkezetét teljesen nem ismerve, azt hitték, hogy C
Tehát ha megnézzük akkor C,O,H-bõl áll, és amit írtál, hogy a cukrokból többféle létezik a C miatt van.
Ha van egy olyan C atom, amihez kapcsolódik 4 különbözõ másik ligandum (atom v molekula), akkor ebbõl a C atomból 2 féle létezik, melyek egymásnak tükörképei (királis), mint a két tenyér:
Ez csak akkor történik meg, ha a C-hez 4 különbözõ szubsztituens kapcsolódik, ha ezek közül 2 egyforma, akkor úgy összeforgatható a két C, hogy azok egymást fedik, tehát nem tükörképek.
A cukrokban is van ilyen C-atom amelyik királis:
http://t2.gstatic.com/images?q=tbn:7MKGzF1J4GvWLM:http://fr.wikivisual.com/images/5/53/D_et_l_glucose.png&t=1
Amint a képbõl is látszik, egymás tükörképe a két molekula, ugyanolyan atomok alkotják, ugyanolyan kapcsolódási sorrendben, de más térszerkezetben.Az egyiket D (dexter-jobb) a másikat L (leveus-bal) glükóznak nevezik, az alapján, hogy a királis C-atom az ún. Fischer-projekciós ábrán merre áll.
dixitque deus fiat lux
#6
upsz, ezt nem ide akartam, hanem másik, bocsánat <#shakehead>
#5
Ezek az alfa béta konformációk ha jól emlékszem, és valóban máshogy törik a fényt, és emiatt másmilyenek. Valamelyik glükózfajtából a természetben talán nincs is, de már nem emlékszek, hogy az alfa, vagy a béta változatból. Szerintem a kutatás legelejének a 10.es kémia könyv pont jó, onnan meg tudod érteni, hogy molekuláris szinten mi a különbség a két molekula között.
#4
Egy átfogó kémia könyvet tudnátok ajánlani nekem?
#3
fú én azt szeretném kérdezni hogy:
régebben olvastam valahol, hogy az ember (meg a szén alapú életformák a földön) csak az olyan tápanyagot (cukrot azthiszem) tud(nak) lebontani, ami bizonyos szögben töri meg a fényt... van ami jobbra, van ami balra és az egyiket nem tudja (valami alfa;béta valami volt a neve)
de nemtudom már hogy egyáltalán hogy kezdjek neki ennek az írásnak a felkutatásának, mert nagyon kulcsszót se tudok. errõl te tudsz valamit vagy vakvágányon vagyok?
régebben olvastam valahol, hogy az ember (meg a szén alapú életformák a földön) csak az olyan tápanyagot (cukrot azthiszem) tud(nak) lebontani, ami bizonyos szögben töri meg a fényt... van ami jobbra, van ami balra és az egyiket nem tudja (valami alfa;béta valami volt a neve)
de nemtudom már hogy egyáltalán hogy kezdjek neki ennek az írásnak a felkutatásának, mert nagyon kulcsszót se tudok. errõl te tudsz valamit vagy vakvágányon vagyok?
#2
elsõként legyen:
A kolera toxin patomechanizmusa
A Vibrio cholerae egy Gram-negatív baktérium, amely egy toxin hatású fehérje komplex kiválasztására képes (tehát exotoxin), ez a kolera toxin (Ctx). A Ctx felelõs a kolera tüneteiért és patomechanizmusának ismeretében választ adhatunk a tünetek kialakulásának miértjére.
A patomechanizmus ismeretéhez, elõször a Ctx szerkezetét kell szemügyre venni.
Az alábbi képen a Ctx sematikus szerkezete látható (PDB_ID:1xtc):

A képen látható piros (CtxA-1) és narancssárga (CtxA-2)fehérjerészlet alkotja az toxin A részét (CtxA) és a kék a B részét (CtxB). A CtxB 5 egyforma (homológ) fehérjerészbõl áll, tehát egy pentamer és egy gyûrût alkot, amelybe beleilleszkedik a CtxA-2 α-hélix része (narancssárga), a két szerkezetet egymással másodlagos kölcsönhatások révén kapcsolódnak, kovalens kölcsönhatások révén nem.
A CtxA α-hélix részét (CtxA-2, narancssárga) és a késõbbiekben enzimatikus szerepû CtxA-1 részt egy cisztein-cisztein (Cys-Cys) diszulfid híd köti össze, amely a meglelõ körülmények között elszakad, szabaddá téve az enzimatikus CtxA-2 részt.

Amint az a kísérletekbõl kiderült a CtxB pentamer szerepe, hogy a sejtbe való bejutást megvalósítsa. Ez úgy történhet meg, hogy kapcsolódok a bélhámsejtek membránjában lévõ, ún. GM1 gangliozid receptorokhoz, melynek hatására a teljes fehérje komplexet bekebelezi a sejt. A kialakult membránnal körülvett struktúrát endoszómának hívjuk. Az endoszóma ezután a Golgi-készülékbe szállítódik, ahol a sejt saját fehérjéi felszakítják a CtxA-1 és CtXA-2 fehérjeláncok közötti Cys(187)-Cys(199) diszulfid kötést, ezáltal szabaddá téve az enzimatikus funkcióval bíró CtxA-1 részt.

Ezek után a CtxA-1 rész a citoplazmába jut, ahol egy másik fehérje (ún. ADP-ribolizációs faktor 6) szabaddá teszi a CtxA-1 aktív centrumát, melynek hatására az képes lesz megkötni egy NAD (Nikotinsavamind-Adenin-Dinukleotid) molekulát. Így a CtxA-1-NAD komplex kötõdik egy membránban lévõ másik fehérjéhez, egy adenilát-ciklázhoz, melnyek feladata cAMP (ciklikus-AdenozinMonoFoszfát) termelése, adott inger esetén, mely normális körülmények között a membránban lévõ G-fehérjéktõl jön. Viszont ebben az esetben a CtxA-1-NAD komplex ADP-ribozilációt hajt végre az adenilát-ciklázon, amely annyit jelent, hogy a CtXA-1-NAD komplex NAD részébõl egy ADP-ribozil részt kapcsol az adenilát-ciklázhoz, ennek eredményeként az adenilát-cikláz folyamatosan elkezd cAMP-ot termelni.
A cAMP másodlagos hírvivõként szolgál általában a sejtekben, a felszaporodása egyáltalán nem rendeltetésszerû. Jelen esetben a létrejövõ hatalmas mennyiségû cAMP egy elektrolit kiáramlást fog okozni a bélhámsejtekbõl a bél lumenbe, amely víz kiáramlását vonja maga után. Ez összességében hasmenést eredményez és dehidratációhoz vezet, amely a kolera elsõ számú tünete és haláloka.
A kolera toxin patomechanizmusa
A Vibrio cholerae egy Gram-negatív baktérium, amely egy toxin hatású fehérje komplex kiválasztására képes (tehát exotoxin), ez a kolera toxin (Ctx). A Ctx felelõs a kolera tüneteiért és patomechanizmusának ismeretében választ adhatunk a tünetek kialakulásának miértjére.
A patomechanizmus ismeretéhez, elõször a Ctx szerkezetét kell szemügyre venni.
Az alábbi képen a Ctx sematikus szerkezete látható (PDB_ID:1xtc):
A képen látható piros (CtxA-1) és narancssárga (CtxA-2)fehérjerészlet alkotja az toxin A részét (CtxA) és a kék a B részét (CtxB). A CtxB 5 egyforma (homológ) fehérjerészbõl áll, tehát egy pentamer és egy gyûrût alkot, amelybe beleilleszkedik a CtxA-2 α-hélix része (narancssárga), a két szerkezetet egymással másodlagos kölcsönhatások révén kapcsolódnak, kovalens kölcsönhatások révén nem.
A CtxA α-hélix részét (CtxA-2, narancssárga) és a késõbbiekben enzimatikus szerepû CtxA-1 részt egy cisztein-cisztein (Cys-Cys) diszulfid híd köti össze, amely a meglelõ körülmények között elszakad, szabaddá téve az enzimatikus CtxA-2 részt.
Amint az a kísérletekbõl kiderült a CtxB pentamer szerepe, hogy a sejtbe való bejutást megvalósítsa. Ez úgy történhet meg, hogy kapcsolódok a bélhámsejtek membránjában lévõ, ún. GM1 gangliozid receptorokhoz, melynek hatására a teljes fehérje komplexet bekebelezi a sejt. A kialakult membránnal körülvett struktúrát endoszómának hívjuk. Az endoszóma ezután a Golgi-készülékbe szállítódik, ahol a sejt saját fehérjéi felszakítják a CtxA-1 és CtXA-2 fehérjeláncok közötti Cys(187)-Cys(199) diszulfid kötést, ezáltal szabaddá téve az enzimatikus funkcióval bíró CtxA-1 részt.
Ezek után a CtxA-1 rész a citoplazmába jut, ahol egy másik fehérje (ún. ADP-ribolizációs faktor 6) szabaddá teszi a CtxA-1 aktív centrumát, melynek hatására az képes lesz megkötni egy NAD (Nikotinsavamind-Adenin-Dinukleotid) molekulát. Így a CtxA-1-NAD komplex kötõdik egy membránban lévõ másik fehérjéhez, egy adenilát-ciklázhoz, melnyek feladata cAMP (ciklikus-AdenozinMonoFoszfát) termelése, adott inger esetén, mely normális körülmények között a membránban lévõ G-fehérjéktõl jön. Viszont ebben az esetben a CtxA-1-NAD komplex ADP-ribozilációt hajt végre az adenilát-ciklázon, amely annyit jelent, hogy a CtXA-1-NAD komplex NAD részébõl egy ADP-ribozil részt kapcsol az adenilát-ciklázhoz, ennek eredményeként az adenilát-cikláz folyamatosan elkezd cAMP-ot termelni.
A cAMP másodlagos hírvivõként szolgál általában a sejtekben, a felszaporodása egyáltalán nem rendeltetésszerû. Jelen esetben a létrejövõ hatalmas mennyiségû cAMP egy elektrolit kiáramlást fog okozni a bélhámsejtekbõl a bél lumenbe, amely víz kiáramlását vonja maga után. Ez összességében hasmenést eredményez és dehidratációhoz vezet, amely a kolera elsõ számú tünete és haláloka.
dixitque deus fiat lux
#1
Üdvözlet minden kedves témalátogatónak ! {:
A Biokémia téma célja, hogy minden érdeklõdõ számára lehetséges legyen a biokémiáról és annak társdiszciplináiról (molekuláris biológia, sejtbiológia, bioinformatika, etc.) a fórum keretein belül megvalósuló társalgása.
Idõközönként írni fogok bõvebben az általam érdekesnek tartott témákról, amelyekrõl majd szívesen várom véleményeteket, hozzászólásaitokat és esetleges kérdéseiteket.
Bármilyen témához kapcsolódó kérdést/megnyilvánulást szívesen várunk!
A Biokémia téma célja, hogy minden érdeklõdõ számára lehetséges legyen a biokémiáról és annak társdiszciplináiról (molekuláris biológia, sejtbiológia, bioinformatika, etc.) a fórum keretein belül megvalósuló társalgása.
Idõközönként írni fogok bõvebben az általam érdekesnek tartott témákról, amelyekrõl majd szívesen várom véleményeteket, hozzászólásaitokat és esetleges kérdéseiteket.
Bármilyen témához kapcsolódó kérdést/megnyilvánulást szívesen várunk!
dixitque deus fiat lux
← ElőzőOldal 4 / 4